Ancient Foodways: Zooarchaeology and Paleoethnobotany
Note – We are currently beta testing this textbook. If you are a student who has been assigned this reading, we would appreciate any feedback you may have. After finishing the chapter, please go to https://forms.gle/3QDm5QYi4ft4wbnXA to complete a short survey.
Steven R. Kuehn, M.S., Illinois State Archaeological Survey
Rebecca K. Albert, M.A., University of California, Santa Barbara
Emily Johnson, M.A., University of California, Santa Barbara
Learning Objectives
-
Define and describe zooarchaeology and paleoethnobotany
-
Outline the primary methods and techniques used in faunal and floral analysis
-
Discuss how zooarchaeological and paleoethnobotanical data provide information on past human behavior and the interrelationship between people and their environment
Zooarchaeology and Paleoethnobotany
Animal and plant remains found at archaeological sites provide important insight on past human behavior and how humans interacted with their environment. The evidence obtained can tell us about past dietary practices, what environments were like in the past, how and when plants and animals were first domesticated, foraging strategies, and how plant and animal resources were used for medicines, tools, and ritual items. Analysis of past animal and plant remains represent two specialized fields in archaeological research.
is the study of animal (zoology) remains from prehistoric and historic archaeological sites. The overall goal of zooarchaeological research is to develop a better understanding of how past peoples interacted with their environment, specifically with animals (fauna). Zooarchaeology is an interdisciplinary field of study that combines zoology and archaeology, but with an emphasis on the latter. Fields such as ecology, biology, osteology, , , and various others also contribute to zooarchaeological studies. Zooarchaeological or are a reflection of past human behavior and living strategies. Analyzing animal remains from archaeological deposits can provide a wealth of information on how past peoples lived (e.g., Russell 2012), not just their behavior but also on the environment in which they lived at the time.
is the study of how people (ethno) used plants (botany) in the past (paleo), or how people interacted with and impacted their environment through the use, manipulation, and consumption of plants. Paleoethnobotany can be used to understand what plants people ate, how people were acquiring plant foods, how people prepared plants for consumption, the origins of agriculture, or what the environment was like at the site, to name a few. Fields such as botany, ecology, agronomy, and soil science also greatly inform paleoethnobotanical studies.
Special Topics: Taxonomic Classification
All biological organisms are scientifically ranked and placed in defined categories in order to group together those sharing similar traits or common ancestry. These taxonomic ranks are hierarchical in structure, meaning that those higher up in the rank are more generally similar, while those lower in rank are more closely related. The scientific names used in are agreed upon by the scientific community (the International Commission on Zoological Nomenclature), so that there is mutual understanding between researchers and guidelines for creating and refining taxonomic ranks. The two most common terms used, genus and species, are written in italics and represent closely related species.
This table displays the taxonomic classification for the coyote and common sunflower:
|
Rank |
Animal |
Plant |
|
Kingdom |
Animalia |
Plantae |
|
Phylum (Division in plants) |
Chordata |
Tracheophyta |
|
Class |
Mammalia |
Magnoliopsida |
|
Order |
Carnivora |
Asterales |
|
Family |
Canidae |
Asteraceae |
|
Genus |
Canis |
Helianthus |
|
Species |
latrans |
annuus |
|
Common name |
coyote |
common sunflower |
An animal closely related to the coyote, such as a red fox, would be in the same Kingdom-through-Family ranks, but would differ by genus and species (red fox is Vulpes vulpes). In scientific research and reporting, authors regularly use the scientific names of animals and plants because common names can vary considerably, causing confusion. The Latin names used in taxonomic classification are accepted and understood world-wide.
At its core, archaeology consists of the analysis and interpretation of left behind by past peoples. Faunal and utilized by prehistoric and historic peoples represent specialized material culture obtained from the living biotic environment (often termed ), and as such are often subsumed under the category of . The data obtained during floral and faunal analysis represents only the initial step used to address research questions in archaeological studies. In order to gather significant information from faunal and floral remains, it is necessary to examine in some detail the methods and techniques employed.
Zooarchaeology and Faunal Analysis
Zooarchaeological faunal analysis focuses on the study of preserved faunal or animal remains recovered from archaeological sites. Typically, the preserved remains consist of the hard parts of an animal such as bones, teeth, antler, shell, and scales. Depending on various taphonomic factors, a representative sample of animal remains may preserve over time, forming a zooarchaeological or faunal assemblage for a particular site. Soft items (e.g., hair, skin, muscle, marrow, etc.) only rarely preserve in the archaeological record, in settings such as unusually anoxic wet (e.g., peat bog) or arid (e.g., dry cave) conditions. During analysis, each faunal remain or specimen is examined individually and a series of characteristic traits and conditions recorded. This initial step focuses on and taxonomic identification; essentially, identifying the part of the animal as well as the type of animal represented by each specimen. (See Special Topics: Taxonomic Classification, below.) The information obtained is used to develop a zooarchaeological profile of the assemblage, which can then be used to address research questions regarding the human behavior that created the assemblage.
Comparative Specimens and Reference Guides
The analysis of zooarchaeological remains begins through specimen recognition and identification. Each piece of bone or shell is examined individually and compared to known specimens, either in a or in . A faunal comparative collection consists of elements from individual animals, all properly cleaned and labeled, that can be used to assist in the identification of typically fragmented or incomplete zooarchaeological remains. Reference manuals consist of books, journal articles, and similar publications that provide detailed illustrations (usually drawings, but also photographs) of skeletal elements that can be used for identification. In general, comparative collections are preferable to reference manuals; it is often difficult to obtain the precision necessary to accurately recognize small, distinct traits visible on skeletal elements through drawings or photographs. In addition, many skeletal remains can be quite small, making them difficult to illustrate. Finally, no reference manuals exist that contain detailed illustrations of every bone from every animal found in a given region.
Animal Remains and Basic Osteology
In order to recognize the variety of faunal remains present in an archaeological assemblage, it is necessary to have a basic familiarity with a diverse array of animal skeletons. The animal kingdom can be divided into and . Vertebrates have backbones or spinal columns, and at least some internal skeletal elements; they include mammals, birds, reptiles, amphibians, and fish. Invertebrates, in contrast, lack a spinal or vertebral column. Common invertebrates include mollusks (bivalves, gastropods, squids, octopi), arthropods (insects, arachnids, crustaceans), annelids (earthworms, leeches), and cnidarians (jellyfish, corals, sea anemones). Other than some mollusk and arthropod remains, Invertebrate remains rarely occur in the zooarchaeological record, so the primary focus is on the analysis of Vertebrate skeletal remains.
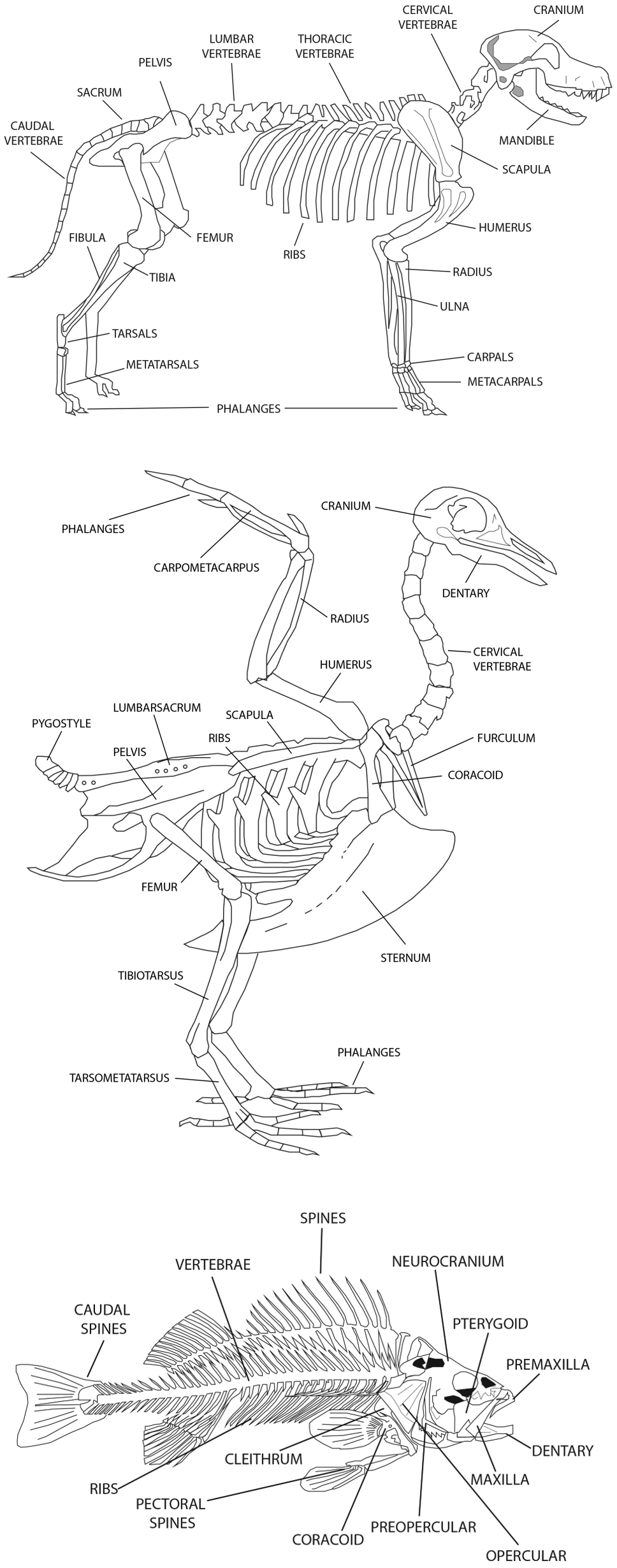
The typical Vertebrate skeleton is comprised of axial and appendicular elements. The consists of the skull (cranium, mandibles, teeth, hyoid) and rib cage (ribs, sternum, vertebrae). The includes the bones of the fore and hind limbs, the shoulder or pectoral girdle, and the hip or pelvic girdle. The number and character of the different axial and appendicular bones varies between taxa, so zooarchaeologists need to be familiar with the general osteological structures of and variations between mammals, birds, reptiles, amphibians, and fish (Figure 1). Each element differs in some respect between animals, with the differences more pronounced between animals that are not closely related. For example, forelimb bones are similar between dogs and foxes, but very different between dogs and beavers. Some animals, such as turtles and fish, have very distinctive skeletal elements that are easily recognized, but differences are still found between the same bones of different species (e.g., catfish bones are easily distinguished from those of freshwater bass). These differences can be used to make class, genus, and species-level classifications between specimens.
Mollusk or shellfish remains are some of the few invertebrate remains typically encountered in a zooarchaeological assemblage. Both bivalves and gastropods (univalves) have hard shells with distinct characteristics (e.g., shape, thickness, color, size and orientation of teeth, presence of exterior bumps or ridges) that can be used for identification. Even when working with fragmentary pieces of shell, examination of multiple characteristics can often be used to distinguish between different species. Whether working with vertebrate or invertebrates remains, however, often the poor condition of the specimens prevents identification to a specific element or distinct taxon. Faunal remains should never be assigned a level of identification that is not wholly supported by the available evidence.
Specimen Identification and Analysis
As part of the analysis, the following data are recorded for each piece of bone: element; side of the body (when applicable); portion or section of the element present; the age of the animal; and taxonomic classification, to the most precise level permissible. For age, an approximation generally equivalent to juvenile or adult is used, based on epiphyseal closure of the long bones, tooth eruption, occlusal wear on the chewing surfaces of the teeth, or similar traits (O’Connor 2000:80-97). (See Special Topics: Aging and Sexing Skeletal Elements, below.) Mollusk shells are treated in a similar manner, with element, portion, and taxon recorded for each specimen.
Special Topics: Aging and Sexing Skeletal Elements
Animal skeletons grow and change throughout the life of the animal. With careful observation, it is possible to recognize and note these changes and use this information to determine the approximate age of the animal at the time of death. The most common of these is tooth eruption. In mammals, for example, deciduous or milk teeth erupt from the gumline first, only to be pushed out and replaced by the permanent or adult teeth. This process occurs at a regular rate, so that it is possible to roughly determine an individual’s age based on the eruption and growth of the teeth present. Adult teeth, once fully erupted, remain with the animal through the rest of their life, but do wear down through use over time. As teeth grind together and chew up food items, the bumps and ridges on the occlusal or chewing surface of the teeth wear down. With some variation, this occlusal wear can be used to determine age. Diet can affect the results (e.g., eating coarse foodstuffs like grasses, for example, can wear teeth down faster), but in general greater occlusal wear is found in older individuals.
Some skeletal elements are made up of smaller pieces of bones, and the fusion of these pieces can also reflect age. As juveniles, mammal skulls are comprised of a series of smaller bones that fuse together as adults. Examining the sutures, where the smaller bones fuse together, can provide evidence of age. Appendicular limb or long bones also change during development. In juveniles, long bones consist of a bony diaphysis and two epiphyses (one on each end) that are separated by cartilage. As the bones grow, the cartilage disappears and the ends of the diaphysis fuse with the two epiphyses, forming a complete, intact bone. Depending on how much fusion has occurred, this can be used to age the element. Each long bone element fuses at a different rate, so each bone can be used to determine specific ages until the animal reaches adulthood. With experience, an analyst can also recognize muscle-attachment wear, arthritis, and other physical signs typically associated with aged individuals.
Skeletal elements are constant within a given species, but for some animals there are differences between males and females. This is an example of sexual dimorphism, where two sexes of the same species display different characteristics. In certain animals, for example, males have a baculum or penis bone, an element not found in females of that species. In some cervid (deer, elk) taxa, males grow antlers while females do not. Thus, if you find a deer cranium with antlers attached, you can recognize it as a male. In other animals such as caribou, however, both males and females grow antlers, although those of the males tend to be larger and more robust. In other animals, the size and shape of horns and antlers can be used to differentiate males from females.
Other skeletal elements may also differ between the sexes, but these differences are not always mutually exclusive. Elements in males are typically larger and more robust than the same elements in females. For example, canine teeth in male carnivores are usually larger than those in female carnivores, but a large female could have elements larger than those of a small male. This problem is compounded when examining fragmentary specimens, which are most common in an archaeological faunal assemblage. Identifying these traits or finding a baculum, for instance , would allow the analyst to recognize male or female elements in a zooarchaeological assemblage. In general, relatively few zooarchaeological remains can be securely identified to sex.
Each faunal specimen is also examined for evidence of non-cultural and cultural . Non-cultural modification, reflecting alterations caused by non-human agents, includes such things as weathering, rodent and carnivore gnawing, and similar taphonomic effects (see Chapter #Traces-Environment#; Lyman 1994). These modifications can impact bone preservation and identifiability; understanding the nature of these impacts aids in accurately documenting and interpreting faunal assemblages. Cultural modifications include those resulting from direct and indirect human activity. Possible cultural modifications include burned or unburned condition; presence and types of butchery marks; and modification relating to the manufacture of bone or shell tools, ornaments, and similar items. Butchery marks include cut marks, fractures, splinters, and other damage caused by carcass processing (e.g., Binford 1981:87-181). Bones, antler, shell, and other animal parts also served as raw materials for a wide array of implements and items used in daily and ceremonial activities. Modified faunal remains can be recognized by cut, scrape, and/or polish marks that resulted from impacts that changed the original shape of the element.
are a unique situation, as they can result from internal (e.g., disease) or external (e.g., trauma) factors, and in the case of the latter can be caused by human behavior (Baker and Brothwell 1980; Bartosiewicz 2013). Animal husbandry, or the domestication and breeding of certain taxa, can also result in changes to animal bone that can be recognized in the zooarchaeological record (O’Connor 2000:147-159).
Special Topics: Microfaunal Identification
Most skeletal remains are visible to the naked eye and can be observed and studied simply by looking at them. Some smaller remains, such as fish scales or rodent bones, can be examined more easily using a magnifying lens or loupe. Certain faunal remains, such as insect parts, are difficult if not impossible to distinguish without visual enhancement. Magnification can also make it easier to identify butchery marks, evidence of tool manufacture, and pathologies.
If identification of remains is so challenging, then is it necessary to do it? In truth, many small fragments or crumbs of bone are often unidentifiable to element or taxon and provide little if any substantial information. Other remains, however, can provide a wealth of knowledge for the observant analyst. Small rodent teeth, for example, can be difficult to identify unless examined under magnification, but they are exceptional environmental and climatic indicators. Rodents represent approximately 40% of all mammalian species on Earth and occur naturally on every continent except Antarctica. They have relatively short life spans, occur in a wide range of biomes, and are sensitive to changes in climate and vegetation. This information can be particularly useful if a scientist wishes to know more about paleoenvironmental conditions and changes over time. As such, putting extra effort into the study and analysis of rodent teeth can yield great rewards.
Recovery Methods, Sampling, and Context
The composition of a faunal assemblage does not reflect a perfect representation of all animal remains present at an archaeological site. Rather, it is a biased sample. Not all faunal material preserves over time; acidic soil conditions, for example, can result in the destruction of more fragile animal remains. In addition, recovery techniques (e.g., hand collection, screening, ) can impact the assemblage composition. Flotation through fine-grained mesh will undoubtedly recover more microfaunal remains than screening through one-quarter inch mesh, for example. Finally, it is rare that an entire archaeological site is completely excavated. As such, only the faunal material present in those areas investigated will ever make it to the zooarchaeologist for analysis.
is likewise an important consideration. It is typically assumed that the majority of faunal material recovered from an archaeological site represents the remains of animals used by the site inhabitants. Some bone and shell recovered may be , meaning that it was deposited before or after human habitation of the site. In addition, some animal remains are , and represent animals that live in or near human habitations but are not directly associated with human activity. For example, in any given backyard there may be mice, voles, snakes, frogs, snails, and other animals unnoticed by the people living in the adjacent house.
Special Topics: Indirect Evidence of Human-Animal Interaction
In addition to bone, shell, teeth, and other faunal remains, indirect evidence can be used to recognize the presence of different animals at prehistoric and historic archaeological sites. Rock art paintings and carvings, for example, often portray various animals that were important to the human inhabitants. Gnaw marks found on recovered bones can indicate the presence of dogs or rodents, even if their physical remains do not occur in a zooarchaeological assemblage. Animal tracks, although rare, are on occasion found at archaeological sites and can likewise demonstrate the presence of certain animals. Dog and cat paw prints, for example, have been found impressed on roofing tiles at sites from the Roman Empire. Material culture relating to animal domestication, such as bridle and tack hardware from horses, can be used to recognize the presence of horses at a site even if no bones are recovered. Finding a pet collar or discarded bird cage in a historic privy could likewise indicate that the residents of the adjacent home once kept a variety of pets.
Insect nests, such as those from mud daubers, are occasionally found among the remnants of burned prehistoric structures in the Southeastern United States and demonstrate the presence of insect pests in human settlements. Paleofeces (e.g., dehydrated fecal remains, typically found in arid or anerobic conditions) are one uncommon type of indirect evidence found at archaeological sites. Both human and dog paleofeces were found at several sites in the American Bottom region of Illinois. Analysis of the faunal remains contained in the dog paleofeces provided direct evidence for dog diet, and showed that the dogs ate various fish, amphibians, and small mammals. These examples represent only a small sample of the instances in which indirect evidence provide insight on human-animal interaction in the archaeological record.
Paleoethnobotany and Floral Analysis
Paleoethnobotany is usually split into two methods of analysis based on the size and type of plant remains being studied: macrobotanical and microbotanical analysis. analysis deals with plant remains such as carbonized seeds and plant parts you can see with your eyes or the aid of a low-powered microscope. analysis deals with plant remains that you can only see with the aid of a high-powered microscope, such as pollen, starch, and phytoliths. is a semicrystalline carbohydrate produced by plants, and is helpful for plant identification because of morphological (shape and size) variation between different families and genera of plants (Messner 2011). are made of silica (like glass, or sand) and help give plants structure by forming in between cell walls when plants uptake silica from groundwater. As with starch, phytoliths can be identified to plant family or genera based on their unique shape and size. Starches are found in carbohydrate-rich plants such as beans, tubers, maize, and cereal grains, while phytoliths are found in greater quantities in the grass family (Piperno 2006). In fact, many grasses feel rough to the touch because of phytoliths. is a fine powder that plants make when they reproduce. Plant and tree pollen is then dispersed by the wind and settles into soil or on top of bodies of water, such as lakes, where it falls to the bottom and accumulates into the sediment over time. Archaeologists can then take core samples of the soil to investigate changes in pollen type and abundance throughout time. Pollen morphology is also unique to families and genera.
This section will broadly cover a few of the sampling, processing, and analytical methods used by paleoethnobotanists to learn about plant use in the past. While macro- and micro-botanical data can be used together to answer similar social and environmental questions, the ways in which each type of botanical data is recovered and processed can be very different.
Considering the environment and conditions at a site can help archaeologists to determine the suitability of certain sampling strategies. As with zooarchaeological remains, sampling strategies are often determined or guided by the unique taphonomy at an archaeological site. For example, botanical remains in archaeological contexts can be affected by the pH of the soil, and whether the botanical sample has been carbonized, waterlogged, or desiccated (completely dried out). Botanical remains that have been carbonized or desiccated tend to preserve in archaeological soils much better than samples that have not. On the opposite side of the spectrum, samples that have been waterlogged, or continually in water protected from the effects of oxygen, also tend to preserve very well. However, once removed from water, these samples can deteriorate quickly. Botanical remains in more acidic soils risk being dissolved at a faster rate, especially if they are not carbonized or desiccated. All of these factors guide what method archaeologists choose to recover botanical remains.
Sampling Methods
Paleoethnobotanical analysis first begins with sampling from a field site; depending on whether an analyst will be doing macrobotanical or microbotanical recovery, these samples can range from soil, residues on artifacts, or dental calculus. Macrobotanical data are derived almost entirely from plant remains found in soil samples (except for the cases where botanical remains are desiccated or preserved in waterlogged contexts), while microbotanical data can be derived from remains isolated from all three sampling methods.
Soil Sampling
Macrobotanical analysis begins with sampling soil both across an archaeological site and within individual features or units. Important factors to consider when collecting soil samples include: 1) What questions researchers want to ask from the data, 2) The context of the site and its features, and 3) the archaeologist’s time, resources, or personnel constraints. But how much soil or residue do you need for a sample, and how does it differ based on the analyses someone wants to do?
Sampling across a site generally takes the form of one or a combination of the four following methods: sampling, sampling, sampling, and sampling (Guedes and Spengler 2014). Generally, blanket sampling (also called full coverage or total sampling) is the preferred sampling method because soil samples are taken from all units and individual features. This reduces the possible bias introduced by other sampling methods (Pearsall 2000). For a further discussion of how factors like time, personnel, person power, and site size might dictate one of the other sampling strategies, see Guedes and Spengler (2014).
In addition, archaeologists must also consider how best to sample within a feature or stratigraphic level. The three most common strategies include: bulk sampling (soil is collected from a single layer within a single context); pinch sampling (for large cultural layers or levels of construction fill, a small amount of soil is collected from various spots and combined into one sample); and column sampling (soil is taken in a line down a single column at each unique stratigraphic level). Unlike site-wide sampling methods, in which there is usually only one strategy in place, for features and contexts it is common to use all three feature-sampling methods listed above.
For macrobotanical (e.g, wood, charcoal, seeds) analysis, the standard practice is to take 10 L of soil for each context sampled (D’Alpoim et al. 2014). This amount of soil is large enough to be representative of the context as a whole. If a researcher wants to answer questions that also require microbotanical analysis, a subsample of soil from the 10 L should be taken at the same time so that both the macro and microbotanical data from the site can be fully integrated. Microbotanical analysis only requires a small amount of soil—roughly 100-300 mL (or half a cup).
Other Sampling Methods
Other common sources for collecting microbotanical data include residues on artifacts and dental calculus. Residues include carbonized food residues from the inside of ceramic sherds, absorbed residues from the fabric of a ceramic sherd, or residues from the cutting edges of lithics, among others. Residues from ceramic sherds can tell archaeologists what people were cooking or storing in ceramic vessels, while residues from lithics can tell archaeologists what people were cutting with these tools. These residues are typically collected in the lab using a sanitized sonicating toothbrush. Dental calculus is mineralized plaque that accumulates in layers on teeth and can be collected from both animal and human remains using a sanitized dental pick or sonicating toothbrush in a lab setting. Microbotanical analysis of dental calculus can inform paleoethnobotanists about what a person was eating directly, unlike the previous methods mentioned, which are only secondary sources of information about diet or how people were using plants in their day-to-day life. Pollen data are recovered by taking a soil core from lake sediment and sampling each stratigraphic level; pollen is used to reconstruct ancient environments.
Processing Soil for Macrobotanical Remains
The process for recovering macrobotanical remains from soil is called flotation. While there are several different methods of flotation, the principle behind the process is to separate the dense material in the sediment (the soil matrix itself, rocks, bone, stone tool flakes, etc.) from the light material (wood charcoal, carbonized and modern plant seeds, plant parts, etc.). These light and heavy fractions are separated by adding the soil sample to water and agitating to 1) dissolve the soil into solution, 2) let the heavy fraction sink, and 3) let the light fraction float. The light fraction is skimmed off of the top and collected in a small piece of chiffon gauze that allows only water to pass through (Figure 2). The remaining heavy fraction is collected in a 2mm window screen mesh. After drying both the light and heavy fractions (out of the sun to avoid heat-induced popping or stress on botanical material; this may take 2-3 days), the heavy fraction is sorted to ensure no plant material remains. Any remaining botanical material is added to the dry light fraction and labeled and sealed in plastic bags for future analysis.
Special Topics: Different Types of Flotation and How Archaeologists Use Them
Flotation is an important step in macrobotanical analysis that paleoethnobotanists use to separate macrobotanical remains such as seeds and wood charcoal from other soil components. While the principle behind flotation is simple—botanical remains will float because they are lighter than water, and bone, stone, pottery, and lithics will sink because they are heavier—there are several ways to go about floating your soil samples depending on the size of the samples, access to water, budget, and other factors.

is the simplest method of flotation, and it can be done just about anywhere. As the name suggests, with bucket flotation an archaeologist adds a small portion of the soil sample to a bucket, adds water, and gently stirs with their hands to break up the dirt and encourage it to go into solution. Next, they slowly decant the water and botanical remains floating on top into another bucket lined with chiffon to catch all of the botanical material. More water is added to the bucket to dissolve more soil and force more botanical remains to float, and the process is repeated until no more botanical remains float to the surface. The heavy fraction left in the bucket is then saved in window screen mesh to look at later for other archaeological material. The pros of bucket flotation are that it requires minimal equipment that is readily available in the field and uses much less water than other methods of flotation. However, it can be a very tedious process if soil samples are large in volume or there are hundreds of samples to process, and it can be difficult to break up soils with a heavy clay content. The next most popular method of flotation is (Shell Mound Archaeological Project), named after the project that created it. This style of flotation requires the construction of a simple machine (Figure 2) that forces water to rise from below into a large tank or bin and pushes the light botanical material to the top of the tank where it can be skimmed off the top in the same manner as in bucket flotation. SMAP machines allow archaeologists to process large volumes of soil and can be inexpensive to make but require some expertise to build and cannot recycle water in areas where it is scarce. Lastly, a machine runs in the same way as a SMAP machine and can similarly process large volumes of soil but allows water to be recycled. However, it is the most expensive method to build.
Each method of flotation has its benefits and drawbacks, and the machine that works best for a project will ultimately be decided by the specific context of the site, soil, budget, and available materials.
Analysis of Macrobotanical Remains
Once back in the laboratory, analysis of macrobotanical remains requires only a few simple tools. A geological sieve is necessary to separate the light fraction into different size classes both to simplify the analysis and ensure that small seeds are not lost within large pieces of grass or fine silt sediment. The most common sieve sizes are 2 mm, 1 mm, 0.5 mm, and finally the pan, which catches the remaining material, but this varies by region and by researcher. For example, some projects use 2 mm, 1.4 mm, 0.7 mm, and pan, while some regions without many small seeds skip the 1.4 mm fraction entirely.
A scale is used to weigh the entire light fraction as well as each individual size class from the geological sieves. This allows the researcher to calculate statistics such as densities and ratios that are valuable in answering archaeological questions (see 5. Paleoethnobotanical Data for more information on common statistical approaches to macrobotanical data). Finally, a low-powered stereo (also called dissection) microscope allows analysts to better see small seeds and unique morphologies necessary for identification and later analysis.
Wood is one of the most ubiquitous botanical remains a paleoethnobotanist will come across in either the field or the laboratory due to its abundant use as a source of fuel and shelter. Wood is primarily encountered as wood charcoal, having been preserved by fire through incomplete burning—either accidentally or intentionally. In rarer cases, at sites with extreme weather conditions a paleoethnobotanist may encounter wood samples that are waterlogged or desiccated. Because wood charcoal is often highly fragmented and it would be impossible to know whether the wood in your sample is from one branch or ten, it is only ever weighed rather than counted. In addition, data for wood charcoal is only recorded at the 2 mm fraction and above since these are representative of the entire sample; the small amounts of charcoal remaining in the other fractions are negligible.
Seeds are the primary material that paleoethnobotanists use to answer anthropological questions regarding past peoples’ foodways, environment, economy, and more. All seeds 1mm and above, both whole and broken, are counted and weighed. Generally, seeds below 1 mm in size are only counted if they are whole, though this may vary by project. Seeds below 1mm are not weighed because they are too light to register on the scale so archaeologists only record count data.
When identifying seeds, it is important to identify distinct morphological features such as size, general shape, surface texture, appearance of the seed fractured vs whole, and more. Much of this knowledge will come from analyzing samples with a skilled graduate student or professor, as well as comparative reference collections both online, physical, and in book form. Popular online reference manuals include: Digital Plant Atlas, the website for the Ohio State University Department of Horticulture and Crop Science, and the Searchable Seed Image and Illustration Gallery, among others. Popular books that have useful images and descriptions of key family characteristics include Seed Identification Manual (Martin and Barkley 2004) and Botanical Macro-remains (Schoch et al. 1988).
Analysis of Microbotanical Remains
After samples are collected, they are chemically processed according to the type of analysis. For example, starch remains are more delicate than phytoliths or pollen, so the same method cannot be used to extract all types of microremains. Chemical processing often involves the separation of materials, often by using specific chemicals to dissolve parts of the sample while leaving others intact. can also be used. Just like standard soil flotation, chemical floatation uses differences in material densities to separate the microbotanical remains from their matrix. This process involves chemicals other than water so that specific densities can be achieved to separate the microremains. Methods for processing these samples can vary greatly, and each lab that analyzes microbotanical samples has a standardized method for each sample type.
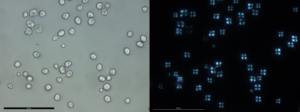
After samples are chemically processed, drops of the isolated microbotanical sample are pipetted onto a slide and mixed with a mounting medium for microscope analysis and identification of diagnostic microbotanical remains. These slides are then analyzed under a high-powered microscope with polarizing- or colored-light filters. Polarizing filters allow for the identification of starches because under polarizing light they have a distinctive cross shape (Figure 3). Colored filters can increase the contrast between the background of the field of view and the microbotanical remain being viewed.
Identification to the level of species or genus usually requires recognizing unique combinations of shape, features, and size. For phytoliths (Figure 4), these shapes include rondel, cross, bilobate, polylobate, bulliform, and hair cells, with different shaped and sized phytoliths coming from different parts of the plant in some occasions (Madella et al. 2005; Piperno 2006). For starches, shapes can include lenticular, bell, elliptical, or oval, with surface features such as fissures, margins, lamellae, or hila (Messner 2011). Like macrobotanical and zooarchaeological analysis, the identification of these diagnostic features is aided through the use of a comparative collection, reference manuals, or online reference websites, and the constraints of these different tools for macrobotanical analysis are the same for microbotanical analysis. For commonly utilized reference manuals and online databases for microbotanical analysis, please see the For Further Information section below.
Zooarchaeological Data
Quantitative Data, NISP, and MNI
Following specimen identification, it is necessary to quantify or count the faunal remains in such a way that useful, meaningful data are collected. are used to measure the relative abundance of each taxa, so that zooarchaeologists can determine what animals were being used by humans, how important each animal was in comparison to other animals used, and what animal representation might reveal about past environmental conditions. The information recorded can be of varying usefulness when analyzing a faunal assemblage. Certain basic data are typically recorded for all faunal specimens; some other types of data can be calculated based on the information obtained. These are referred to as primary and , respectively (see Grayson 1984; Lyman 2008; Reitz and Wing 1999:142-238).
The most common recorded is the . The NISP documents the number of specimens identified to each element for a given taxon. For example, if an assemblage contains three rabbit molars, two rabbit vertebrae, nine raccoon rib fragments, and seven raccoon humerus pieces, the NISP for rabbit would be five and for raccoon would be 16. The taxon does not have to be specific, however; if six long bone shaft fragments are classified as unidentifiable large mammal, then the NISP for large-sized mammal would be six. It should also be noted that NISP counts recognize partial elements, as well as side of the body, age, size, and other factors. Suppose that the same assemblage contains the following deer bones: one left femur, proximal end only; one left femur, distal end only; two right femora, both complete; one femur, mid shaft portion only. The deer NISP is five, because each specimen is counted separately even though some of the femur fragments may be from the same element. The NISP is the basic unit of data from which most other quantitative measures in faunal analysis are derived.
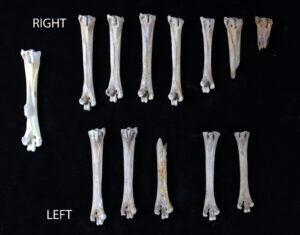
One drawback of the NISP is that it can, on the surface, overestimate the presence and therefore the relative importance of a particular taxon. Using the above example, there are more raccoon bones (NISP=16) than rabbit (NISP=5) or deer (NISP=5) bones in the assemblage, which would seem to suggest that raccoons were used to a greater extent than rabbits or deer. The NISP does not, however, reflect the number of individuals represented in the assemblage, only the number of specimens or pieces of bone. To compare individuals, the analyst must calculate the . MNI calculations are based on repeating elements from the same taxon (Figure 5). If an assemblage contained four right ulnae and two left ulnae identifiable as muskrat, the MNI for muskrat would be four. Minimally, at least four individual muskrats must be represented, as only one right ulna is found in a single animal. There could be as many as six muskrats represented, if the two lefts were from totally separate animals, but this cannot be determined with certainty. In general, paired elements are the most useful in determining MNI; bones that are similar and/or occur in relatively large numbers, such as ribs, are less suitable and less likely to provide significant results.
The MNI is usually only calculated for specimens that can be identified to the genus and/or species level, but some exceptions are made on occasion (see Reitz and Wing 1999:198-199). Unfortunately, this means that MNI estimates are not made for general classification categories (e.g., “indeterminate fish” or “indeterminate mammal”). As a result, potentially significant information on faunal representation may be lost if only the MNI is used. If, for example, an assemblage contains 500 fish bones but none are identifiable to the genus level, then none will be included if the analyst only focuses on MNI results. MNI can also be impacted by preservation to a greater extent than NISP. If bones are poorly preserved, it is less likely that they can be classified to the genus/species level. In most faunal analyses, both NISP counts and MNI determinations are included, so that complementary lines of information are available.
Secondary Data
In addition to NISP and MNI, there is an array of other data that can be gathered from a faunal assemblage. Most of these, however, are drawn from or derivative of NISP or MNI calculations, or do not provide information that reflects past human behavior. Specimen weight (used to calculate ) may be recorded, for example, but the results have more to do with the initial size and robustness of the bone itself and do not necessarily reflect relative dietary importance. For example, one deer bone might weigh as much as several hundred small fish bones; that does not mean, however, that deer were necessarily a more important food item, but the raw weight values would seem to support such an argument. In contrast, the NISP and MNI counts would suggest that fish were of greater dietary value than deer. Lyman (2008:84-113) and Reitz and Wing (1999:222-231) provide excellent summaries of the pros and cons of various types of primary and secondary zooarchaeological data (see also Grayson 1984).
Paleoethnobotanical Data
After paleoethnobotanical data are quantified (counted and/or weighed), additional statistical methods can be employed to examine patterns within the data (Marston 2014). The simplest way to report botanical data is through absolute counts, which are the raw counts for each individual taxon identified. Other descriptive statistics frequently used are:
- Density calculations: seed or charcoal counts/weights are compared against the volume of the original soil sample taken in the field to provide comparability between all samples taken at a site.
- Proportions/Percentages: these calculations are used to standardize the count of one taxon versus another category of taxon. For example, a comparison of the ratio between maize kernels and maize cupules at a site can tell you where ancient people were processing dried maize (high cupule ratio) versus using the kernels for cooking (high kernel ratio).
- Ubiquity: presence/absence data are used to calculate the percentage of samples in which a given taxon appears. For example, if you have 10 samples and maize is found in 6 of them, the ubiquity value of maize is 0.60 and tells us maize is present in 60% of samples.
Other types of statistics in paleoethnobotanical reports often include Z-scores (data are transformed into standard deviations around a population mean), diversity indices (calculated based on the relative values of taxa to indicate homo- or hetero-geneity of a sample), and more complicated multivariate calculations such as cluster analysis, principal components analysis, and correspondence analysis (see Smith 2014 for discussion and examples of multivariate statistics in paleoethnobotany). Once calculated, these statistics can be presented in a number of ways to highlight patterns or trends revealed by your analyses.
Ancient Foodways: What Can Zooarchaeology and Paleoethnobotany Reveal?
Faunal and floral remains from archaeological sites can provide a wealth of information on past human behavior and environmental interaction. Some of the more common areas of study are addressed here, but on-going research is discovering ever more applications for zooarchaeological and paleoethnobotanical data.
Dietary Patterns
Zooarchaeological and paleoethnobotanical material provide direct evidence for prehistoric and historic diet. The faunal and floral remains recovered do not reflect all of the food refuse discarded in the past, but they do provide a robust sample of what people were eating. This information provides important insight on how animals and plants were processed for consumption, and how human dietary strategies changed over time. Not all faunal and floral remains recovered, however, represent food refuse; this is addressed in detail below.
Floral remains recovered demonstrate what sorts of wild plants were gathered, what domesticated crops were grown, and how important various taxa were to the site inhabitants. Similarly, the faunal remains recognized reveal what wild animals were procured, what domesticated animals were utilized by the site inhabitants, and the importance of different animals to the inhabitants. The proportional amounts of animal and plant remains illustrates their relative significance in the diet. By examining assemblages from different sites and different time periods, it is possible to see how dietary strategies varied between regions and changed over time.
In addition, recognizing patterns in what people were eating and how they were processing those types of foods can inform archaeologists about cuisine and the foodways (culture surrounding food) at archaeological sites. Combining dietary analysis with ceramic and lithic use-wear analyses, for example, enables archaeologists to employ multiple lines of evidence towards a more complete understanding of food consumption as well as the social ramifications of dining, feasting, and gender roles (e.g., Kooiman 2018; Twiss 2012).
Foraging Strategies
The types of animals and plants recovered at an archaeological site can provide insight on the hunting and foraging practices employed by the site inhabitants. Certain faunal and floral species are available or abundant only at certain times of the year, and human foragers will often optimize their efforts by focusing on these taxa only when they are likely to yield high returns. For example, nut trees such as oaks only produce ripe acorns in the fall, so foraging groups might schedule their seasonal travels so that they are close to oak forests during the autumn months. Likewise, these same mobile foragers might plan to camp near large lakes or rivers in the spring, when spawning fish are in the shallows and more easily caught. By recognizing the time of year when different taxa are available, archaeologists can often determine the (season of occupation) for a site.
The relative abundance of different animal and plant remains also reflects foraging behavior. Some hunter-gatherer groups focus their efforts on a select range of large animals (e.g., big-game hunters), although most employ a broad-based subsistence strategy in which they procure a wide array of animals and plants to fulfill their dietary needs. Other groups may adjust their foraging strategies on a seasonal basis in order to take advantage of available, abundant resources. Agricultural communities, for example, put considerable effort into farming during the spring planting and fall harvest seasons, but during the other seasons supplement their diets through hunting, fishing, and foraging.
Paleoenvironmental Reconstruction
Different types of plants prefer different environmental conditions, so the presence, absence, disappearance, or appearance of plant species through time at a single site may indicate changes in environment such as drought, desertification, increased precipitation, or changing temperatures. These changes in local habitat can also be indicative of human actions that impact the environment such as overgrazing, tree harvesting, agriculture, environmental management, and many others (e.g., Asouti and Austin 2005). One method of recognizing these changes is through analysis of pollen cores, which can reveal distinct changes in environmental conditions over long periods of time (See Special Topics: Pollen Coring and Environmental Interactions, below.)
Special Topics: Pollen Coring and Environmental Interactions
How can archaeologists, environmental scientists, and geographers learn more about the environment surrounding lakes, and in effect the actions of the people occupying the areas around this lake? The analysis of pollen, known as palynology, can help us understand these environmental changes on a local level—palynologists collect sediment core samples from the floors of bodies of water and analyze the pollen, the male gamete of plants, within those samples. Pollen, unlike other microbotanical remains, can usually be identified to the species level, and therefore can give palynologists a detailed picture of the surrounding plant environment. Pollen also collects in layers of sediments at the bottom of bodies of water chronologically. Pollen at the bottom of a core sample will be the oldest, and pollen at the top of a core sample will be the youngest (MacDonald and Edwards 1991). Cores collected from these bodies of water can vary in length depending on what periods of time the analysts are interested in, and how fast these layers accumulate locally.
After these cores are collected, palynologists sample these cores for radiocarbon or Accelerated Mass Spectrometry (AMS) dating and process the remaining samples to dissolve the sediment, usually with a chemical like hydrofluoric acid (HF). After processing, palynologists identify and quantify all the pollen from the samples in order to understand the surrounding plant community. From these dated samples, palynologists can then model the changes in the plant environment surrounding the body of water throughout time, and what factors may be at play. For example, if there is an increase in pollen from agricultural crops, such as maize, in tandem with a decrease in different types of tree pollens, that would indicate that there was deforestation for agricultural purposes (Alenius et al. 2012).
Similar information can be derived from analysis of the different types of faunal represented at a site. Depending on the sensitivity of the taxa, it is possible to reconstruct what a habitat was like during the period a site was occupied. Many animal species flourish only within restricted habitats, and these taxa can be particularly useful when examining environmental changes over time.
Plant and Animal Domestication
Tracking changes in seed size (from smaller to larger) across time is one indication that a plant may be undergoing domestication. The recovery of non-local plants in archaeological contexts, such as maize in Eastern North America (e.g., Larson et al. 2014), can be evidence of domestication (as well as interregional exchange, as discussed below). Similar changes are often seen among domesticated animals. Domesticated animals typically exhibit physical traits that reflect breeding efforts to increase meat production or reduce aggression. It can take many generations for these changes to manifest themselves skeletally, but they can be recognized in the archaeological record over time.
Butchery Practices
Close examination of animal bones may reveal evidence of butchery, such as cut and chop marks, but the bones themselves also provide insight on how carcasses are butchered and processed. First, the location and types of marks identified are direct evidence of the specific manner in which animal carcasses were reduced in size and portioned for further processing. Butchery marks are commonly found near limb joints and muscle attachment points, which reflects the removal of meat-rich limb portions. Other types of cuts, such as those found in areas with little muscle, may represent cuts associated with hide or pelt removal.
The bones themselves—and the absence of some bones—are likewise indicative of choices made during the butchering process. If an animal is killed at a great distance from the habitation site, for instance, hunters may choose to leave low-value lower limb or cranial portions at the kill site. As a result, lower leg and skull bones may be under-represented in the zooarchaeological record of the habitation, but much more common at kill locations. A large percentage of high-value bones (those supporting large amounts of meat, or choice cuts) in an assemblage may indicate the differential distribution of meat resources, possibly reflecting tribute, feasting, or allocation of foodstuffs based on status. Finally, heavily fragmented appendicular bones can also be indicative of marrow extraction and bone grease manufacture.
Trade and Resource Movement
The recovery of faunal or floral remains from species not typically found in an area can provide evidence of trade, population mobility, and similar movement of people and resources. For example, marine whelk shells and shark teeth are occasionally found at prehistoric sites across the Eastern United States, far inland from coastal areas. These items were likely transported, either directly or indirectly, by traders and travelers moving between these areas. The recovery of tropical plant species in temperate zones (e.g., maize, as noted above) reflects not only interregional trade but evidence of domestication as well. Exotic items were undoubtedly highly valued and served to encourage further contact and commerce. It is likely that many other goods, most of them perishable and not recognized in the archaeological record, were also exchanged between neighboring groups.
Non-Dietary Use of Faunal and Floral Material
The non-dietary use of faunal and floral resources is often overlooked, with most research focused on dietary strategies and paleo-environmental reconstructions. A wide range of non-dietary uses are recognized (e.g., Russell 2012), many more than can be addressed in this chapter. A sample of these uses is presented here.
Plant resources were widely used for non-dietary purposes but are often difficult to identify because they typically do not preserve well in the archaeological record. Wood was used extensively for construction projects, tools, bowls, utensils, weapons, canoes, and numerous other items in addition to being burned for fuel. Grass, reeds, plant fiber, and pine needles were used to create cordage, woven basketry, mats, and rugs (e.g., Nichols et al. 2000). Storage containers were manufactured from dried gourds such as Lagenaria siceraria (bottle gourd).
The ritual and medicinal use of plants was similarly significant. Tobacco (Nicotiana tabacum), peyote (Lophophora williamsii), ayahuasca (Banisteriopsis caapi), and other mind-altering substances such as opium poppy (Papaver somniferum) are derived from plants. Various forms of alcohol likewise come from plant products and have mind-altering effects. These items were widely employed by shamans, healers, and ruling elite in a variety of ceremonial and ritual activities (e.g., Morehart et al. 2005; Pauketat et al. 2002).
Many faunal remains similarly represent resources employed in non-dietary roles. Certain items, such as animal hides, sinew, and feathers (for arrow and spear fletching) were likely used for clothing, shoes, and other items but rarely preserve archaeologically. Bone, antler, shell, and other faunal material were fashioned into a variety of items including perforating implements (awls, needles, pins, chisels), flintknapping tools (flakers, billets), weaving and hide-processing tools (shuttles, hide scrapers, beamers), gardening instruments (scapula hoes, digging sticks, antler rakes), gaming pieces (dice, gaming sticks), hunting items (spear points, fish hooks, harpoon heads), and decorations/ornaments (hairpins, beads, combs, pendants). Mussel shell was carved into beads, buttons, gorgets, spoons, and scoops; large marine whelks were transformed into bowls and containers. Turtle shells were similarly made into bowls and rattles.
Faunal remains, both modified and unmodified, were also important as ritual and ceremonial paraphernalia. Small carnivore jaws were often cut and polished, and perhaps functioned as items of sympathetic magic. Bone and shell effigies have also been recovered and may have been used in a similar fashion. Numerous other examples exist, but at times it is difficult to determine the exact function of these items. Bone, antler, shell, and other faunal material modified into wide array of tools, ornaments, decorations, and ritual items.
Summary: Case Studies
Zooarchaeological and paleoethnobotanical remains can provide considerable insight on past human behavior at prehistoric and historic archaeological sites. Much of this information relates to dietary strategies and paleoenvironmental reconstruction, but faunal and floral remains provide significant information on the non-dietary uses of these resources as well. To fully understand how zooarchaeological and paleoethnobotanical data are employed in archaeological research, two case studies are provided.
Special Topics: Case Study, Early Twentieth-Century Dining in East St. Louis, Illinois
Archaeological investigations in the Goose Hill neighborhood, a turn of the century working-class section of East St. Louis, recovered substantial zooarchaeological remains from several different households that allowed for the investigation of differential dietary patterns in an urban residential neighborhood. The information summarized here is taken from Dappert-Coonrod and Kuehn (2017).
The Goose Lake neighborhood in East St. Louis was established shortly after the Civil War, and by the turn of the twentieth century was inhabited primarily by working-class individuals and families, many of whom were employed at the nearby National Stockyards and associated meat-packing plants. In addition to residences, the Goose Lake neighborhood also contained numerous commercial shops and services that supported these residents. Archaeological investigations were conducted in this location as part of the New Mississippi River Bridge Project, with all work overseen by the Illinois State Archaeological Survey (ISAS) of the Prairie Research Institute, University of Illinois at Urbana-Champaign. In addition to the archaeological field and laboratory work, extensive historical and archival research was conducted on this property, in order to better understand the social and cultural context of this historic community.
Zooarchaeological material chosen for analysis was selected from three households: 1) the Mead House, also known as the Maple Hotel or the Davis Boardinghouse, a boardinghouse establishment occupied by a variety of tenants who likely resided for a few months or years; the Benner household, 2) a single family long-term residence owned by Elizabeth Benner, a widow, and her daughter Victoria; and 3) a rental property at 819 Rear Bowman Avenue, characterized as an informal worker’s cottage, and occupied by an ever-changing series of tenants, usually couples or small families. Multiple privy deposits were associated with each household, and they contained relatively large amounts of faunal material discarded by the respective site inhabitants. This allows for a detailed comparison of dietary practices between the occupants of the Mead House, Benner household, and Tenant (or multi-tenant) household.
Cattle, swine, and chicken remains occur in all three assemblages and predominate among specifically identifiable fauna. Mutton (sheep) remains were found in the Mead House assemblage, and eggshell fragments were recovered in the Mead House and Benner assemblages. Fish remains (e.g., carp, brown bullhead) were recognized in all three assemblages, and cottontail rabbit bones were found in the Mead House and Benner assemblages. Other taxa recognized include double-crested cormorant in the Benner household assemblage, turkey in the Mead House assemblage, and graylag (domestic) goose in the Tenant household assemblage. Overall, the composition of the assemblages indicate that the various site inhabitants relied on beef, pork, and chicken (and eggs) as dietary mainstays, but also included other domesticates (e.g., sheep, turkey, goose) and some wild fauna (e.g., rabbit, various fish) in the diet. Dog bones from at least two dogs were found in the Mead House assemblage, suggesting that pets were kept by some of the residents. Rodent bones were also recognized, indicating that urban pests like mice and rats were a problem.
The cuts of meat, particularly beef and pork, represented by the elements recovered provide a more intimate look at dining in the different households. The Mead House inhabitants consumed more steaks, roasts, and hams (as well as chicken and egg dishes) and less soups and stews. At the Benner house, they ate a variety of steaks, roasts, hams, stews, and soups, making use of lesser-quality cuts of meat. The occupants of the Tenant house, however, are more soups and stews than the residents of the Mead House or Benner household. Most of the Tenant assemblage bones were from low-value cuts, particularly beef, suitable mainly for soups and stews. By purchasing cheaper cuts of beef, the Tenant residents were able to buy at least some moderately more expensive pork hams. This indicates that dietary choices were affected, at least to some extent, but the economic means of each household. The residents of the Mead House boardinghouse received at least some higher-cost beef and pork cuts, as part of their room and board. The widow Mrs. Benner was moderately well off financially, it would seem, but still needed to be frugal in her spending. The Tenant household occupants, however, were renters with limited funds available, and this is reflected in their dietary choices.
Interestingly, the serving dishes and other tableware recovered during the archaeological investigations at each property support the patterns observed in the faunal remains. The Mead House boardinghouse dishes included large serving platters for group meals, but also dishware associated with the serving of steaks and roasts. The Benner tableware, in contrast, contained more fashionable dishes, perhaps reflecting the income and status associated with single-family home ownership. The dishes found in the Tenant household privies were of somewhat lower value, but there were also some fancier porcelain dishes comparable to those found at the nearby Benner house. While no single line of evidence is sufficient to answer all questions regarding dietary choices, purchasing power, and socioeconomic status, looking at faunal remains and ceramic dishware together can provide important insight into these research issues.
Special Topics: Case Study, Synthesis of Macro- and Microbotanical Data at the Site of Los Naranjos, Honduras
Los Naranjos is a major multi-component archaeological site in Honduras that lies on the northern border of Lake Yojoya. The site has monumental architecture and sculpture that date to between 1000 and 500 BCE, as well as use of jade and pottery styles that link it to the Gulf Coast Olmec. The inhabitants of the site were thought to have been dependent on maize agriculture as their primary food source by 800 BCE, an assumption that has been challenged by recent paleoethnobotanical work that combines both macrobotanical and microbotanical analyses. This case study, from an article by Morell-Hart et al. (2014), will reveal why it is important to use multiple lines of evidence when considering human relationships with plants in the past.
A lake pollen core taken from previous excavations in the 1980s suggested a meaningful variation in plant pollen over time. Levels from the core corresponding to earlier time periods revealed a high percentage of tree species and no crops such as maize or beans. Progressing through time, researchers interpreted a rise in plants such as Chenopods (the family that contains the popular grain quinoa), maize, and other grasses as evidence of early agriculture. Agricultural intensification was then interpreted later on in time from an increasing percentage of plants from the sunflower family, ragweed, and other plants that like habitats that have been disturbed by human activity. However, the pollen from this core comes primarily from wind-pollinated plants and therefore does not provide a complete picture of plant use, nor does it show direct evidence of plant use at the site of Los Naranjos itself.
To fill in the informational gaps left by the existing pollen record, the authors of this paper analyzed plant data from six kinds of contexts including macrobotanical samples from light fractions, and microbotanical samples taken from the edges of obsidian artifacts used as tools. The samples come from interior surfaces, matrices between surfaces, architectural and pit fill, and a or trash dump.
Maize was recovered by the analysts in several different forms, including as phytoliths, starch granules, and a charred kernel. However, other typically ubiquitous parts of maize (cupule and cob fragments) were not identified. Amaranth and chia seeds were also found, and starch granules of manioc and sweet potato were identified from residues on obsidian tools. Tree crops were important for early agriculturalists, and botanical evidence from the site suggests that people were using palm species for a variety of culinary and non-edible uses. Interestingly, species that are commonly thought of as belonging to the Mesoamerican agricultural suite (beans and squash, primarily) were not evident at the site. Plants with medicinal or cultural uses were identified through both macro- and microbotanical analyses, including tobacco, various grass species, and sunflower species.
The early analysis of the pollen core led researchers to believe that maize agriculture, as well as its two other sisters, beans and squash, were present in the region by 1000 BCE. However, microbotanical analysis of starch granules and phytoliths along with macrobotanical recovery of whole seeds paints a different picture. The recent paleoethnobotanical interpretation reveals that as opposed to widespread intensive agriculture, the residents of Los Naranjos were likely exploiting a wide variety of plants, including seed, root, and tree crops. This new analysis also illustrates the use of plants for non-culinary purposes, notably the use of tobacco which may have been used ritually as well.
While the pollen data were not incorrect, their interpretation of human activity without considering other lines of evidence led to a conclusion that did not show the whole picture of what was happening at the site. Flotation is important for recovering macrobotanical material but is not as directly associated with human activity as microbotanical residues sampled directly from artifacts. Synthesizing a variety of botanical data enriches our understanding of human practices, including non-culinary uses of plants, and complicates our view of the past.
Discussion Questions
- What can animal and plant remains from archaeological sites tell us about past peoples and how they lived?
- Aside from diet, what information about past lifeways and environmental conditions can be determined from animal and plant remains from archaeological sites?
- One important part of archaeological research is the capability to examine how human behavior shifted over time. Provide one example of how changes in daily life might be recognized by comparing different faunal and floral assemblages from two sites from different time periods.
- Zooarchaeological and paleoethnobotanical remains do not always preserve over time. Describe how the absence of certain animal and plant remains might impact our view of life in the past.
- Following up on Question 4, are there ways for archaeologists to fill in knowledge “gaps” in the archaeological record when studying the past? What types of indirect evidence might archaeologists use to obtain more detailed insight into the past? (For example, even if no horse bones were recovered, the presence of horseshoes and metal bridle pieces would indicate horses were present at a site.)
- Faunal and floral resources were often used in ritual and ceremonial activities, as well as for utilitarian or everyday activities. Discuss two examples each of animal and plant resources used in ritual activities. Can you provide a similar example from modern-day life?
- Archaeologists studying animal and plant remains specialize in the identification and analysis of faunal and floral remains. Discuss how knowledge of ecology, zoology, botany, and other fields can provide important insight for interpreting the remains recovered.
About the Authors
 Steven Kuehn is a Zooarchaeologist with the Illinois State Archaeological Survey, a division of the Prairie Research Institute of the University of Illinois at Urbana-Champaign. He has over thirty years of professional experience as an archaeologist, working primarily in the Upper Midwest and Great Lakes Region. Steven has analyzed zooarchaeological remains from Paleoindian through Historic Euroamerican sites in the Midwest, Great Plains, Southeastern United States, and Alaska. Some of his recent research focuses on Paleoindian subsistence behavior, Archaic bison hunting in Minnesota, and the archaeology of domestic dogs in the Midwest. In his free time, Steven enjoys reading, hiking, and playing board games with his family. Steven also writes short stories and mystery fiction, including novels in his Jacob Caine Archaeology Mystery series.
Steven Kuehn is a Zooarchaeologist with the Illinois State Archaeological Survey, a division of the Prairie Research Institute of the University of Illinois at Urbana-Champaign. He has over thirty years of professional experience as an archaeologist, working primarily in the Upper Midwest and Great Lakes Region. Steven has analyzed zooarchaeological remains from Paleoindian through Historic Euroamerican sites in the Midwest, Great Plains, Southeastern United States, and Alaska. Some of his recent research focuses on Paleoindian subsistence behavior, Archaic bison hunting in Minnesota, and the archaeology of domestic dogs in the Midwest. In his free time, Steven enjoys reading, hiking, and playing board games with his family. Steven also writes short stories and mystery fiction, including novels in his Jacob Caine Archaeology Mystery series.
 Rebecca Albert earned her M.A. in Archaeology at the University of California, Santa Barbara in 2021. She was previously the Assistant Coordinator for the Central Coast Information Center, one of nine archaeological and built environment data clearinghouses in the California Historic Resources Information System. Her undergraduate and graduate research focused on microbotanical analysis of carbonized residues from the Upper Great Lakes to gain a deeper understanding of the history of plant domestication in North America, and how these plants may have informed prehistoric foodways. In her free time, Rebecca enjoys quilting, cooking, and playing with her cat, Juliet.
Rebecca Albert earned her M.A. in Archaeology at the University of California, Santa Barbara in 2021. She was previously the Assistant Coordinator for the Central Coast Information Center, one of nine archaeological and built environment data clearinghouses in the California Historic Resources Information System. Her undergraduate and graduate research focused on microbotanical analysis of carbonized residues from the Upper Great Lakes to gain a deeper understanding of the history of plant domestication in North America, and how these plants may have informed prehistoric foodways. In her free time, Rebecca enjoys quilting, cooking, and playing with her cat, Juliet.
 Emily Johnson is a Paleoethnobotanist pursuing her Ph.D. in Anthropology at the University of California, Santa Barbara. She has conducted excavations and/or analyses of archaeological sites in Turkey, Israel, Guatemala, Mexico, and throughout the United States. Her current research is focused on investigating the timing, spread, and development of the nixtamalization process throughout ancient Mesoamerica. When not reading about all things corn, Emily enjoys crafting and playing with her cat, Scout.
Emily Johnson is a Paleoethnobotanist pursuing her Ph.D. in Anthropology at the University of California, Santa Barbara. She has conducted excavations and/or analyses of archaeological sites in Turkey, Israel, Guatemala, Mexico, and throughout the United States. Her current research is focused on investigating the timing, spread, and development of the nixtamalization process throughout ancient Mesoamerica. When not reading about all things corn, Emily enjoys crafting and playing with her cat, Scout.
Further Exploration
Microbotanical reference manuals:
PhytCore DB – phytcore.org – Free with registration online database of photomicrographs of phytoliths, searchable by shape, type of sample, site, etc.
Starch Grain Database – clarissacagnato.weebly.com/starch-grain-database.html – Free online database of starch grains collected from modern samples
Cooked Starch Database – figshare.com/Cooked_Starch_Database/3734335 – free image database of starches from plants that have been cooking various ways. From Henry, Hudson, and Piperno, 2009.
References
Albert, Rebecca K., Susan M. Kooiman, Caitlin C. Clark, and William A. Lovis (2018) Earliest Microbotanical Evidence for Maize in the northern Lake Michigan Basin. American Antiquity 83(2):345-355.
Alenius, Teija, Teemu Mökkönen, and Antti Lahelma (2012) Early Farming in the Northern Boreal Zone: Reassessing the History of Land Use in Southeastern Finland through High-Resolution Pollen Analysis. Geoarchaeology 28(1):1-24.
Asouti, Eleni, and Phil Austin (2005) Reconstructing Woodland Vegetation and Its Exploitation by Past Societies, Based on the Analysis and Interpretation of Archaeological Wood Charcoal Macro-Remains. Environmental Archaeology 10:1–18.
Baker, John, and Don Brothwell (1980) Animal Diseases in Archaeology. Academic Press, New York, New York.
Bartosiewicz, László (2013) Shuffling Nags, Lame Ducks: The Archaeology of Animal Disease. Oxbow Books, Oxford, United Kingdom.
Binford, Lewis R. (1981) Bones: Ancient Men and Modern Myths. Academic Press, Orlando, Florida.
Dappert-Coonrod, Claire P., and Steven R. Kuehn (2017) A Boarder, a Widow, and a Tenant Sit Down for Dinner: Foodway Comparisons in the Goose Hill Neighborhood, East St. Louis, Illinois. Midcontinental Journal of Archaeology 42(1):4-36.
Grayson, Donald K. (1984) Quantitative Zooarchaeology: Topics in the Analysis of Archaeological Faunas. Academic Press, Orlando, Florida.
Guedes, Jade D’Alpoim, and Robert Spengler (2014) Sampling Strategies in Paleoethnobotanical Analysis. In Methods and Theory in Paleoethnobotany, edited by John M. Marston, Jade D’Alpoim Guedes, and Christina Warinner, pp. 77-94. University Press of Colorado, Boulder, Colorado.
Kooiman, Susan M. (2018) A Mutliproxy Analysis of Culinary, Technological, and Environmental Interactions in the Northern Great Lakes Region. Ph.D. Dissertation, Department of Anthropology, Michigan State University, East Lansing, Michigan.
Larson, Greger, Dolores R. Piperno, Robin G. Allaby, Michael D. Purugganan, Leif Andersson, Manuel Arroyo-Kalin, Loukas Barton, Cynthia Climer Vigueira, Tim Denham, Keith Dobney, Andrew N. Doust, Paul Gepts, M. Thomas P. Gilbert, Kristen J. Gremillion, Leilani Lucas, Lewis Lukens, Fiona B. Marshall, Kenneth M. Olsen, J. Chris Pires, Peter J. Richerson, Rafael Rubio de Casas, Oris I. Sanjur, Mark G. Thomas, and Dorian Q. Fuller (2014) Current Perspectives and the Future of Domestication Studies. Proceedings of the National Academy of Sciences 111(17):6139–46.
Lyman, R. Lee (1994) Vertebrate Taphonomy. Cambridge Manuals in Archaeology. Cambridge University Press, Cambridge, United Kingdom.
Lyman, R. Lee (2008) Quantitative Paleozoology. Cambridge University Press, New York.
MacDonald, Glen M., and Kevin J. Edwards (1991) Holocene Palynology: Principles, Population and Community Ecology, Palaeoclimatology. Progress in Physical Geography: Earth and Environment 15(3):261-289.
Madella, Marco, Anne Alexandre, and Terry Ball (2005) International Code for Phytolith Nomenclature 1.0. Annals of Botany 96(2):253-60.
Marston, John M. (2014) Ratios and Simple Statistics in Paleoethnobotanical Analysis: Data Exploration and Hypothesis Testing. In Methods and Theory in Paleoethnobotany, edited by John M. Marston, Jade D’Alpoim Guedes, and Christina Warinner, pp. 163-179. University Press of Colorado, Boulder, Colorado.
Martin, Alexander C., and William D. Barkley (2004). Seed Identification Manual. The Blackburn Press, Caldwell, New Jersey.
Messner, Timothy C. (2011). Acorns and Bitter Roots: Starch Grain Research in the Prehistoric Eastern Woodlands. University of Alabama Press, Tuscaloosa.
Morehart, Christopher T., David L. Lentz, and Keith M. Prufer (2005) Wood of the Gods: The Ritual Use of Pine (Pinus spp.) by the Ancient Lowland Maya. Latin American Antiquity 16(3):255–74.
Morell-Hart, Shanti, Rosemary A. Joyce, and John S. Henderson (2014) Multi-Proxy Analysis of Plant Use at Formative Period Los Naranjos, Honduras. Latin American Antiquity 25(1):65–81.
Nichols, Deborah L., Mary Jane McLaughlin, and Maura Benton (2000) Production Intensification and Regional Specialization: Maguey Fibers and Textiles in the Aztec City-State of Otumba. Ancient Mesoamerica 11(2): 267–291.
O’Connor, Terry (2000) The Archaeology of Animal Bones. Sutton Publishing, Phoenix Mill, United Kingdom.
Pauketat, Timothy R., Lucretia S. Kelly., Gayle J. Fritz, Neal H. Lopinot, Scott Elias, and Eve Hargrave (2002) The Residues of Feasting and Public Ritual at Early Cahokia. American Antiquity 67(2): 257–79.
Pearsall, Deborah M. (2015). Paleoethnobotany: A Handbook of Procedures, 3rd edition. Routledge, England, United Kingdom.
Piperno, D. R. (2006). Phytoliths: A comprehensive guide for archaeologists and paleoecologists. AltaMira Press, Lanham, Maryland.
Reitz, Elizabeth J., and Elizabeth S. Wing (1999) Zooarchaeology. Cambridge Manuals in Archaeology. Cambridge University Press, Cambridge, United Kingdom.
Russell, Nerissa (2012) Social Zooarchaeology: Humans and Animals in Prehistory. Cambridge University Press, New York.
Schoch, Werner H., Barbara Pawlik, and Fritz H. Schweingruber (1988). Botanical macro-remains. Haupt, Switzerland.
Twiss, Katheryn (2012) The Archaeology of Food and Social Diversity. Journal of Archaeological Research 20(4): 357–395.
White, Chantel E., and China P. Shelton (2014) Recovering Macrobotanical Remains: Current Methods and Techniques. In Methods and Theory in Paleoethnobotany, edited by John M. Marston, Jade D’Alpoim Guedes, and Christina Warinner, pp. 95-114. University Press of Colorado, Boulder, Colorado.
The study of animal remains recovered from prehistoric and historic archaeological sites, to better understand how past human-animal interaction.
The study of the transition from the biosphere to the lithosphere, anything that happens to a body from the time of death to the time of discovery.
The domestication and breeding of select animal taxa.
Any and all animal (fauna) remains encountered at archaeological sites.
The study of how people used plants in the past.
The systematic ranking of biological organisms based on hierarchical categories that reflect the sharing of similar traits or common ancestry.
In archaeology, a general term used to refer to all tangible items created, used, and left behind by past cultures; this includes not only artifacts but larger items such as structures and cities.
Any and all plant (flora) remains encountered at archaeological sites.
Inclusive term used to represent faunal and floral material obtained from the biotic environment and used by prehistoric and historic peoples.
Subdiscipline of archaeology that focuses on the relationship between past societies and the environment in which they lived; typically includes zooarchaeology, paleoethnobotany, and geoarcheology.
In zooarchaeology and bioarchaeology, term used to refer to a single or individual bone, tooth, scale, scute, valve, or similar faunal remain.
An assemblage of correctly identified, labeled, and complete specimens that can be used to assist in the identification of unknown, typically fragmented, specimens.
A published book, journal, or similar document or record that contains detailed information—typically illustrations—that can be used for specimen identification.
Animals with a spinal column or backbone, and at least some internal skeletal elements; refers to the subphylum Vertebrata, within the phylum Chordata in the animal kingdom (kingdom Animalia).
Animals lacking an internal vertebral column or backbone; this term refers to all animals not found in the subphylum Vertebrata.
In the Vertebrate skeleton, elements associated with the skull, spinal column, and rib cage.
In the Vertebrate skeleton, elements associated with the fore and hind limbs, the pectoral or shoulder girdle, and the pelvic or hip girdle.
Any impact or activity that results in the physical alteration of a faunal (or floral) remain; cultural modifications are those caused directly or indirectly by human activity, such as butchery or tool manufacture; non-cultural modifications are those resulting from non-human or natural activity, such as carnivore or rodent gnawing, weathering, and similar taphonomic processes.
The study of pathologies (trauma, disease, and congenital defects) found in ancient human and animal populations.
Small animal remains that are difficult to discern with the naked eye; artificial magnification is typically needed for accurate identification.
The process of using water to separate soil into its components that are heavier and lighter than water.
The specific location in which an artifact is found, and by association its relationship to other artifacts and ecofacts recovered from the same specific location.
An item found in archaeological context which was deposited before or after the human habitation or activity directly associated with a particular component, and as such does not reflect the activities that created the site.
In zooarchaeology, refers to all organisms living in or around humans and human habitations that are not typically considered dietary items or faunal resources; in general, the humans neither benefit nor are harmed by the presence of these animals.
Plant remains you can see with your bare eyes or a low-powered microscope, including seeds and wood charcoal.
Plant remains that you can only see with a high-powered microscope, including starch, phytoliths, and pollen.
A semicrystalline carbohydrate produced by plants.
Rigid structures made of silica, formed in plant tissues to provide support.
A fine powder that plants make when they reproduce.
A sampling strategy also known as "full coverage" or "total sampling," which takes samples from all units and individual features.
A sampling strategy also known as 'judgmental sampling,' where samples are taken primarily from units and features that are thought to be of interest.
A sampling strategy in which samples are selected through chance, or through arbitrary selection.
A sampling strategy through which samples are selected at regular intervals across the site or within the unit.
A method of flotation done by hand, in which soil is simply dissolved into a bucket so that any botanical remains will float to the surface.
A method of flotation that uses a simple machine.
A flotation machine
The use of chemicals of differing densities to separate microbotanical remains from their matrix.
Information with a value that can be measured or counted (quantified), and have a distinct numeric value; contrasts with qualitative data, which refer to information that observed, characterized, described, and recorded but not quantified.
Information derived from primary data using indices and quantitative techniques; in contrast to primary data, secondary data are typically analytical products rather than based on direct observations or measurements.
In zooarchaeology and paleoethnobotany, information obtained directly from the faunal or floral remains undergoing analysis; primary data are obtained during the initial stage of the analysis, and the results can be replicated during subsequent analysis.
The number of identifiable faunal remains associated with a recognized taxonomic group in a zooarchaeological assemblage.
The minimum number of individuals represented by the faunal remains identified to a recognized taxonomic group in a zooarchaeological assemblage.
In zooarchaeology, a type of secondary data that uses allometric scaling to estimate relative dietary significance based on comparison of faunal specimen weight.
Determination of what season (or seasons) of the year in which a site was occupied, based on the recovery of faunal and floral remains from species present or particularly abundant (e.g., migratory waterfowl) during a particular season, or by the presence of select seasonal indicators (e.g., growth lines in deciduous trees, or antlers in certain cervids).
An accumulation of garbage, usually discarded in a hole in the ground
